A number of genes have been reported to be involved in these steps of biofilm formation. Some of these genes were selected for evaluation of their susceptibility to gene expression inhibition by CCG-203592 using a real time RTPCR approach. The genes down-regulated or up-regulated by CCG-203592 are involved in biofilm VE-822 formation at different stages of biofilm formation. The icaADBC operon encodes enzymes involved in biosynthesis of polysaccharide intercellular adhesin or polymeric N-acetyl-glucosamine that plays important roles in biofilm formation. Deletion of the ica locus significantly decreased S. aureus biofilm formation. Downregulation of icaA could decrease production of PIA/PNAG, leading to reduction of biofilm formation. Interestingly, icaA was up-regulated during ML phase, but down-regulated at 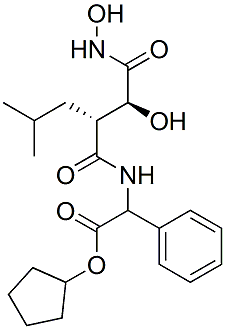 S phase. The net outcome of the effect of CCG-203592 on icaA could result from the combined effect of the dynamic changes of gene expression. The dltABCD operon encodes four proteins responsible for esterification of teichoic acids with D-alanine. Deficiency in dltA results in a stronger negative net charge on the CP-358774 citations bacterial cell surface and defects in the initial binding of bacteria to the surface in biofilm formation. Down-regulation of dltD in the same operon could have similar effects. Autolysin altA is a major peptidoglycan hydrolase that cleaves newly synthesized peptidoglycan components before they are incorporated into the cell wall. Primary attachment of bacteria to surfaces is impaired in altA null mutants. SPA gene was consistently down-regulated by CCG-203592 in all three phases tested. SPA is able to induce cell aggregation and biofilm formation. sdrD is one of the microbial surface components recognizing adhesive matrix molecules that play important roles in mediating bacteria adhesion to host tissues and forming biofilm though the exact function of sdrD is unkown. sspB encodes a cysteine protease that is regulated by agr system. Inactivating sspC which is an inhibitor of sspB, enhances the attachment of bacteria to solid surfaces and biofilm formation, suggesting that sspB has positive effects on biofilm formation. SigB is an alternative sigma factor that regulates a large regulon and inactivating SigB decreases biofilm formation by S. aureus and increases RNAIII level. RNAIII is a component of the agr quorum-sensing system which regulates gene expression in response to outside signals. Inhibition of agr system is important for biofilm development and agr also mediates biofilm dispersal. The influence of agr system on biofilm development is multifaceted and complicated, depending on experimental conditions. Hla was shown to be required for S. aureus biofilm formation and deficiency in Hla caused defects in biofilm formation. Taken together, down-regulating the above genes could negatively impact biofilm formation. On the other hand, psma operon encodes four short PSMa peptides. Deletion of psma causes defects in formation of biofilm channels and biofilm detachment and regrowth which suggested that PSMs are important for biofilm maturation and detachment. Lack of PSMs led to increased biofilm volume and thickness. The lrg operon is responsible for inhibition of murein hydrolase activity of the CidA protein. Mutant inactivating LrgAB operon exhibits increased biofilm adherence and matrix-associated eDNA, and forms biofilm with reduced biomass and defective structures compared.
S phase. The net outcome of the effect of CCG-203592 on icaA could result from the combined effect of the dynamic changes of gene expression. The dltABCD operon encodes four proteins responsible for esterification of teichoic acids with D-alanine. Deficiency in dltA results in a stronger negative net charge on the CP-358774 citations bacterial cell surface and defects in the initial binding of bacteria to the surface in biofilm formation. Down-regulation of dltD in the same operon could have similar effects. Autolysin altA is a major peptidoglycan hydrolase that cleaves newly synthesized peptidoglycan components before they are incorporated into the cell wall. Primary attachment of bacteria to surfaces is impaired in altA null mutants. SPA gene was consistently down-regulated by CCG-203592 in all three phases tested. SPA is able to induce cell aggregation and biofilm formation. sdrD is one of the microbial surface components recognizing adhesive matrix molecules that play important roles in mediating bacteria adhesion to host tissues and forming biofilm though the exact function of sdrD is unkown. sspB encodes a cysteine protease that is regulated by agr system. Inactivating sspC which is an inhibitor of sspB, enhances the attachment of bacteria to solid surfaces and biofilm formation, suggesting that sspB has positive effects on biofilm formation. SigB is an alternative sigma factor that regulates a large regulon and inactivating SigB decreases biofilm formation by S. aureus and increases RNAIII level. RNAIII is a component of the agr quorum-sensing system which regulates gene expression in response to outside signals. Inhibition of agr system is important for biofilm development and agr also mediates biofilm dispersal. The influence of agr system on biofilm development is multifaceted and complicated, depending on experimental conditions. Hla was shown to be required for S. aureus biofilm formation and deficiency in Hla caused defects in biofilm formation. Taken together, down-regulating the above genes could negatively impact biofilm formation. On the other hand, psma operon encodes four short PSMa peptides. Deletion of psma causes defects in formation of biofilm channels and biofilm detachment and regrowth which suggested that PSMs are important for biofilm maturation and detachment. Lack of PSMs led to increased biofilm volume and thickness. The lrg operon is responsible for inhibition of murein hydrolase activity of the CidA protein. Mutant inactivating LrgAB operon exhibits increased biofilm adherence and matrix-associated eDNA, and forms biofilm with reduced biomass and defective structures compared.
Month: August 2019
In vitro studies indicated OSCS can activate the contact system with dependant activation of the kinin-kallekrein system
Interestingly, CidA was up-regulated Carfilzomib during ML and LL phases which could generate similar phenotype as down-regulating lrg. However, mutations in both lrg and CidA caused aberrant biofilm maturation, suggesting that imbalance in their gene expression could CHIR-99021 disrupt biofilm development. These effects of CCG-203592 may increase biofilm formation, which could be outweighed by the effects of down-regulation of other genes by CCG-203592. As a result, the combined effect of all the affected genes by CCG-203592 may produce net decrease of biofilm formation. Interestingly, CCG-203592 decreased the RNAIII level slightly, suggesting that up-regulation of RNAIII level by decreased SigB and CodY level was compensated by changes in other genes that may also regulate RNAIII level. CodY is another global gene regulator that represses agr and icaADBC operon. Inhibition of CodY could have different effects on biofilm formation. Inactivating CodY could enhance biofilm formation in S. aurues strain SA564 and UAMS-1, but reduce biofilm formation in high-biofilmproducing S. arueus isolate S30. More genes were affected by CCG-203592 at stationary phase than at growing phases. We also observed that an analog of CCG203592 changed expression of more genes at stationary phase than at growing phases in GAS. It was well known that expression patterns of many genes are changed at different growth phases. For example, depletion of glucose and change of pH after a long period of culture at stationary phase could impact the gene expression of agr system. As a result, it is possible that CCG203592 has different impacts on gene expression at different growth phases. In order to understand the mechanism of action of this novel anti-virulence compound, further studies on the impact of gene expression changes at different growth phases on biofilm formation are needed. Of note, some of the genes that have been down-regulated also play important roles in staphylococcus virulence. SPA, Hla and PSMs are virulence factors and  sspB plays important roles in staphylococcus evasion and resistance to host defense. Based on the gene profile changes by CCG-203592, downregulation of these genes could lead to defects in biofilm formation at different stages and could also lead to diminished virulence. In conclusion, this class of novel anti-virulence compounds demonstrates inhibitory effects on gene expression of multiple S. aureus virulence factors, especially genes known to be involved in biofilm formation, resulting in significant inhibition of biofilm formation. The compounds also inhibit SK gene expression in GAS, suggesting that this class of compounds could target a gene regulatory mechanism that is conserved between GAS and S. aureus. This class of compounds could be a starting point for development of novel anti-microbial agents against multiple pathogens. Oversulfated chondroitin sulfate, a member of the family of glycosaminoglycans which includes, heparin, heparan sulfate, dextran sulfate, chondroitin sulfate A, CSB, CS-C, CS-E and their oversulfated forms, was found to be a major contaminant in heparin during the period of time in 2007�C 2008 with increased heparin adverse events. Clinical symptoms induced by OSCS-contaminated heparin included: hypotension, nausea and shortness of breath within 5 to 10 minutes after intravenous injection of the drug.
sspB plays important roles in staphylococcus evasion and resistance to host defense. Based on the gene profile changes by CCG-203592, downregulation of these genes could lead to defects in biofilm formation at different stages and could also lead to diminished virulence. In conclusion, this class of novel anti-virulence compounds demonstrates inhibitory effects on gene expression of multiple S. aureus virulence factors, especially genes known to be involved in biofilm formation, resulting in significant inhibition of biofilm formation. The compounds also inhibit SK gene expression in GAS, suggesting that this class of compounds could target a gene regulatory mechanism that is conserved between GAS and S. aureus. This class of compounds could be a starting point for development of novel anti-microbial agents against multiple pathogens. Oversulfated chondroitin sulfate, a member of the family of glycosaminoglycans which includes, heparin, heparan sulfate, dextran sulfate, chondroitin sulfate A, CSB, CS-C, CS-E and their oversulfated forms, was found to be a major contaminant in heparin during the period of time in 2007�C 2008 with increased heparin adverse events. Clinical symptoms induced by OSCS-contaminated heparin included: hypotension, nausea and shortness of breath within 5 to 10 minutes after intravenous injection of the drug.
Several signaling pathways have been shown to play a role in PI-induced cytotoxicity structure similar to bortezomib
Recently, in contrast to our speculation, Chauhan et al. reported the effectiveness of MLN9708 to overcome bortezomib resistance. As several mechanisms have been proposed for bortezomib resistance in addition to ?5 subunit mutations, MLN9708 may be effective for such cases. HPDs are expected to compensate for the weak points of bortezomib as well as the second generation PIs described above, because HPDs are non-peptide agents that inhibit all three catalytic subunits of the proteasome with equal kinetics and could be orally bioactive. Moreover, crystal structure analyses indicate that the binding mode is completely different from that of bortezomib and NPI-0052. This ensures the activity of this agent against Nutlin-3 bortezomib-resistant cells, which was experimentally proven in this study, and probably against cells developing the resistance to NPI-0052. Moreover, we have found that oral administration of K-7174 is indeed effective and is not associated with obvious toxicities, including leukocytopenia, in a murine xenograft model. These features provide a rationale 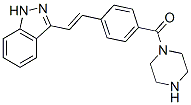 for the clinical translation of HPDs as novel PIs with effectiveness for the Rapamycin mTOR inhibitor treatment of bortezomibresistant patients, a low probability of acquired drug resistance, and flexibility in dosing schedules. In a plethora of in vitro studies it has been extensively demonstrated that inhibition of the proteasome for instance by the tripeptide aldehyd MG-132 or the dipeptide boronate bortezomib selectively kills tumor cells of varying origin. Proteasomal inhibitors also sensitize cells to radio- and chemotherapy and even to apoptosis induced by death receptor ligands. However, as the proteasome targets not only pro-, but also anti-apoptotic proteins, a successful combination therapy requires a successive application of first the apoptosis-inducing agent ensuring the breakdown of anti-apoptotic proteins followed by the PI treatment that then prevents degradation of the generated pro-apoptotic proteins. Nevertheless, bortezomib was the first PI used in clinical trials and approved to treat patients suffering from multiple myeloma or mantle cell lymphoma. Although the new generation of proteasome inhibitors such as salinosporamide and carfilzomib appear to exhibit somewhat different mechanisms of action than bortezomib, central to apoptosis induction by many PIs is certainly the mitochondrial or intrinsic death pathway, as their cytotoxic activity is almost completely abrogated in cells deficient for Bax and Bak. Consistently, a number of studies strongly implicated certain proapoptotic BH3-only proteins in PI-induced apoptosis. For instance, the pro-apoptotic cleavage product of Bid, t-Bid, is degraded by the proteasome and treatment of HeLa cells with MG-132 resulted in accumulation of t-Bid and sensitized the cells to death receptor-induced apoptosis. Also Bik and Bim were found to be upregulated following PI treatment and cells deficient for both or cells in which Bik and Bim were down regulated by RNA interference were refractory to its cytotoxic action. Likewise, different PIs including bortezomib and MG-132 were shown to induce expression of Noxa in several tumor models both at the protein and mRNA level and siRNA-mediated knockdown of Noxa partially rescued various tumor cells from PI-induced apoptosis. Expression of other Bcl-2 family members such as Puma, Bax, Bak, Bcl-2, and Bcl-XL remained mostly unaffected following treatment of different cell lines with PIs.
for the clinical translation of HPDs as novel PIs with effectiveness for the Rapamycin mTOR inhibitor treatment of bortezomibresistant patients, a low probability of acquired drug resistance, and flexibility in dosing schedules. In a plethora of in vitro studies it has been extensively demonstrated that inhibition of the proteasome for instance by the tripeptide aldehyd MG-132 or the dipeptide boronate bortezomib selectively kills tumor cells of varying origin. Proteasomal inhibitors also sensitize cells to radio- and chemotherapy and even to apoptosis induced by death receptor ligands. However, as the proteasome targets not only pro-, but also anti-apoptotic proteins, a successful combination therapy requires a successive application of first the apoptosis-inducing agent ensuring the breakdown of anti-apoptotic proteins followed by the PI treatment that then prevents degradation of the generated pro-apoptotic proteins. Nevertheless, bortezomib was the first PI used in clinical trials and approved to treat patients suffering from multiple myeloma or mantle cell lymphoma. Although the new generation of proteasome inhibitors such as salinosporamide and carfilzomib appear to exhibit somewhat different mechanisms of action than bortezomib, central to apoptosis induction by many PIs is certainly the mitochondrial or intrinsic death pathway, as their cytotoxic activity is almost completely abrogated in cells deficient for Bax and Bak. Consistently, a number of studies strongly implicated certain proapoptotic BH3-only proteins in PI-induced apoptosis. For instance, the pro-apoptotic cleavage product of Bid, t-Bid, is degraded by the proteasome and treatment of HeLa cells with MG-132 resulted in accumulation of t-Bid and sensitized the cells to death receptor-induced apoptosis. Also Bik and Bim were found to be upregulated following PI treatment and cells deficient for both or cells in which Bik and Bim were down regulated by RNA interference were refractory to its cytotoxic action. Likewise, different PIs including bortezomib and MG-132 were shown to induce expression of Noxa in several tumor models both at the protein and mRNA level and siRNA-mediated knockdown of Noxa partially rescued various tumor cells from PI-induced apoptosis. Expression of other Bcl-2 family members such as Puma, Bax, Bak, Bcl-2, and Bcl-XL remained mostly unaffected following treatment of different cell lines with PIs.