Here we show that in contrast with YiiP, ZnT3 forms Labetalol hydrochloride covalent dimers mediated by intermolecular dityrosine bonds. Dityrosine bond formation occurs both spontaneously and induced by oxidative stress. Using site-directed mutagenesis we identified two critical tyrosines in the carboxyterminal of ZnT3, one that prevented and one that increased dimerization. Our studies indicate that covalent tyrosine dimerization of a SLC30A family 4-(Benzyloxy)phenol member modulates its subcellular localization and zinc transport capacity. One modification that could explain formation of covalent dimers regulated by oxidative stress is dityrosine bond formation. Based on the chemistry of dityrosine synthesis, we selected two criteria to identify dityrosine bonds in the 80 kDa ZnT3 species. First, dityrosine formation is induced by oxidative stress and second, is abrogated by tyrosine to phenylalanine mutation of critical residues. Dityrosine formation has been described mainly as a product of oxidative stress and as a normal extracellular posttranslational modification only in a limited group of secreted proteins, such as collagen and elastin. We used crosslinking and sucrose sedimentation to evaluate whether dimer formation in tyrosine mutants occurs spontaneously in the absence of H2O2 as we observed 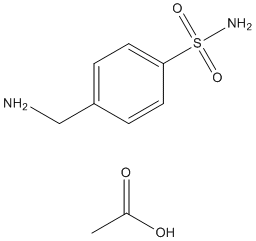 previously for wild type ZnT3. Until now, dimerization of polytopic transmembrane proteins has been shown to occur by covalent and non-covalent interaction mainly through transmembrane domains. Covalent cysteine-based dimer formation has been extensively described for neurotransmitter transporters, such as the dopamine transporter, DAT and the glycine transporter as well as receptors. The fact that dimeric forms of ZnT3 were resistant to reducing agents and increased in response to oxidative stress, lead us to investigate tyrosine-mediated dimerization. Since its discovery in 1959, dityrosine formation has been described as a post-translational modification related with cellular stress and disease. Dityrosine modifications are produced in response to oxidative stress, aging, UV and c irradiation. Increased levels of dityrosine have been found in atheromatous plates, cataracts, acute inflammation, systemic bacterial infection and recently associated with a-synuclein fibrillogenesis and Ab amyloid oligomerization. Only in a limited group of structural proteins of the bacteria cell wall and insect ligaments.
previously for wild type ZnT3. Until now, dimerization of polytopic transmembrane proteins has been shown to occur by covalent and non-covalent interaction mainly through transmembrane domains. Covalent cysteine-based dimer formation has been extensively described for neurotransmitter transporters, such as the dopamine transporter, DAT and the glycine transporter as well as receptors. The fact that dimeric forms of ZnT3 were resistant to reducing agents and increased in response to oxidative stress, lead us to investigate tyrosine-mediated dimerization. Since its discovery in 1959, dityrosine formation has been described as a post-translational modification related with cellular stress and disease. Dityrosine modifications are produced in response to oxidative stress, aging, UV and c irradiation. Increased levels of dityrosine have been found in atheromatous plates, cataracts, acute inflammation, systemic bacterial infection and recently associated with a-synuclein fibrillogenesis and Ab amyloid oligomerization. Only in a limited group of structural proteins of the bacteria cell wall and insect ligaments.
Month: June 2019
Suitable for inferring major demographic changes in a species history
However, species-wide Atropine sulfate samples are not suitable for investigating population processes of divergence, demographic change, and local adaptation. To understand the maintenance of variation within species, and the importance of local selection and demography in natural populations, both within- and amongpopulation sampling are needed. Despite the importance of local population processes to plant evolution, surprisingly little attention has been given to the distribution of DNA sequence diversity among plant populations. Although many studies have used genetic markers to study genetic differentiation among plant populations, so far only a handful have examined DNA sequence diversity within and among natural plant populations. Unlike other molecular marker systems, DNA sequence data provide information about recombination and linkage disequilibrium, which can be highly sensitive to demographic history. Work done to date has demonstrated the need for local sampling to accurately describe patterns of LD, diversity, and the frequency spectrum of polymorphisms in local populations, and shown that even simple demographic processes can better explain observed data than can the assumption of neutral equilibrium. Nonetheless, many of these studies have relied on small samples of loci or groups of candidate genes, neither of which is likely sufficient to capture patterns of genome-wide variation or provide insight into the relative roles of demographic history and selection. Here we present a large-scale population-genetic analysis of sequence diversity in natural populations of Arabidopsis lyrata. A. lyrata is a predominantly self-incompatible, perennial species with a circumpolar distribution across northern and central Europe, Asia, and North America. A. lyrata appears to maintain large, stable populations, particularly in Central Europe, where populations are hypothesized to have served as refugia during the most recent Ice Age. A. lyrata has become a model system for plant molecular population genetics and for investigating local adaptation. For example, divergent selection on trichome production has been found among phenotypically differentiated Swedish A. lyrata populations. Flowering time and floral display also appear to be under strong selection, with large differences in day-length Ginsenoside-F4 requirements between Northern and Southern populations. A. lyrata is 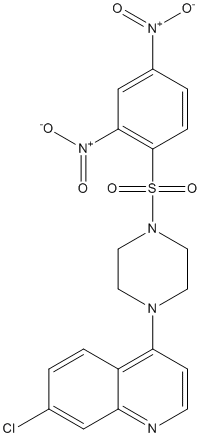 also of great interest because it is a close relative of A.
also of great interest because it is a close relative of A.
It is well posed types to permits shifts in the balance between them
Both activated T helper cells and activated NK cells exhibit signs of being important to SLE pathology, even at the transcriptional level. Perhaps their relative levels define clinically important subtypes of SLE, and if so they might be useful diagnostic markers for this Ginsenoside-F2 disease characteristic. Several 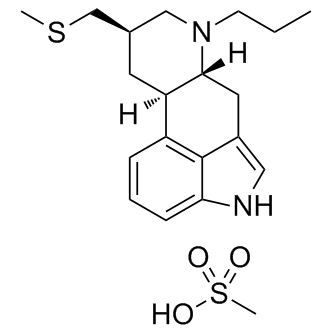 of the cell types that are hypothesized to be particularly important in SLE are correlated with patients�� clinical measurement of disease activity. Activated dendritic cells are efficient antigen presentation machinery, and this cell type is found here to be more abundant in patients with relatively active disease, consistent with correlations between autoantibody blood titer and SLE disease activity. Other immune diseases may be amenable to the methods presented here. One challenge will be the interpretation of predictions in a disease that affects specific organs such as rheumatoid arthritis where there is less evidence of a systemic effect. Deconvolution of biopsies from these patients would have to be interpreted with caution as individual cell types are difficult to isolate from solid tissues for any validation. Moreover, biopsies would contain significant quantities of cells of unknown type. Therefore their signature genes would not be included in the basis matrix for deconvolution. Of course, expanding the arsenal of data on basis cell types would enable thorough analysis of such diverse kinds of samples. Expression deconvolution has several advantages over cell sorting or other traditional methods for quantification of cell species. First, conventional methods deal with at most a handful of different cell types at one time, while expression analysis can simultaneously quantify a much greater number of cell types. Second, deconvolution integrates the partially redundant information of a large number of genes to yield its results; this redundancy is important because it mitigates the contribution of noise inherent in biological measurements and thus Echinatin boosts robustness. Third, post-hoc analysis of expression datasets is relatively rapid, and there are a large and growing number of datasets publicly available. Novel methods of expression analysis like deconvolution have the potential to further tap this important resource for the scientific community. One concern with the application of a system of linear equations such as used here is the mathematical structure of the problem.
of the cell types that are hypothesized to be particularly important in SLE are correlated with patients�� clinical measurement of disease activity. Activated dendritic cells are efficient antigen presentation machinery, and this cell type is found here to be more abundant in patients with relatively active disease, consistent with correlations between autoantibody blood titer and SLE disease activity. Other immune diseases may be amenable to the methods presented here. One challenge will be the interpretation of predictions in a disease that affects specific organs such as rheumatoid arthritis where there is less evidence of a systemic effect. Deconvolution of biopsies from these patients would have to be interpreted with caution as individual cell types are difficult to isolate from solid tissues for any validation. Moreover, biopsies would contain significant quantities of cells of unknown type. Therefore their signature genes would not be included in the basis matrix for deconvolution. Of course, expanding the arsenal of data on basis cell types would enable thorough analysis of such diverse kinds of samples. Expression deconvolution has several advantages over cell sorting or other traditional methods for quantification of cell species. First, conventional methods deal with at most a handful of different cell types at one time, while expression analysis can simultaneously quantify a much greater number of cell types. Second, deconvolution integrates the partially redundant information of a large number of genes to yield its results; this redundancy is important because it mitigates the contribution of noise inherent in biological measurements and thus Echinatin boosts robustness. Third, post-hoc analysis of expression datasets is relatively rapid, and there are a large and growing number of datasets publicly available. Novel methods of expression analysis like deconvolution have the potential to further tap this important resource for the scientific community. One concern with the application of a system of linear equations such as used here is the mathematical structure of the problem.
Consistent with the existence of a regulatory system that constrains the maximum total level
Relatively poor cell type marker but is very valuable when quantifying cell types by the Gentamycin Sulfate methods used here, since it is significantly differentially expressed between many different cell types. This gene is also important here because it is well-established to be induced by type 1 interferons. Among the activated cell types considered here, RSAD2 expression is high in those that are found to be more abundant in SLE patients. Therefore, this gene and the results here based on it support current views that the interferon signature observed in most SLE patients is due to the simultaneous activation of several classes of leukocytes by type 1 interferons and that this class of cytokines is central to the disease. The dynamics of activation appear to differ among cell types. For instance, monocytes constitute approximately the same fraction of blood in all samples Diperodon examined, and the distribution of the proportion of resting to activated monocytes was fairly uniform across its observed range. In contrast, both NK and T helper lymphocytes were essentially fully activated or fully resting. Unlike FACS, the algorithm used here cannot differentiate between a population of a given cell type that are all homogeneously mildly activated and a population of that cell type that consists of a mixture of some resting cells and some strongly activated cells, since the parameters used  to assess activation are merely the total quantity of each gene��s mRNA present in the sample. The monocytes observed here might possess the ability to adopt a mildly activated state, or perhaps they function in discrete activation states like lymphocytes appear to but differ in that the entire population of monocytes need not all be resting or activated in unison. In some cases the level of activation of a patient��s white blood cells was correlated among different cell types. For instance, dendritic cells�� and NK cells�� levels of activation are positively correlated. However, some types of lymphocytes exhibit more complex negative relationships. The maximum level of activated NK cells and T helper cells in SLE patients appears to be constant at approximately 30%. This is clearly seen in most SLE patients, but healthy individuals typically have substantially fewer activated lymphocytes. Although the results presented here offer no direct evidence about the mechanisms by which this pattern occurs.
to assess activation are merely the total quantity of each gene��s mRNA present in the sample. The monocytes observed here might possess the ability to adopt a mildly activated state, or perhaps they function in discrete activation states like lymphocytes appear to but differ in that the entire population of monocytes need not all be resting or activated in unison. In some cases the level of activation of a patient��s white blood cells was correlated among different cell types. For instance, dendritic cells�� and NK cells�� levels of activation are positively correlated. However, some types of lymphocytes exhibit more complex negative relationships. The maximum level of activated NK cells and T helper cells in SLE patients appears to be constant at approximately 30%. This is clearly seen in most SLE patients, but healthy individuals typically have substantially fewer activated lymphocytes. Although the results presented here offer no direct evidence about the mechanisms by which this pattern occurs.