Atorvastatin treatment elicits larger vascular diameter, thus contributing to enhanced regional blood flow perfusion and neuron rescue. So, larger diameters of blood vessels seem to be associated with a better tissue perfusion, protecting neuronal cells from degeneration. Yet, we didn’t observe increased axon regeneration in treated spinal cords compared to control ones. In the same context, Dray’s study suggests that even if some axons benefit from vascular support to accelerate their growth, this support is only transient and limited in time. Tissue protection could also be related to a reduced apoptosis rate within the injured cord. Our data show that BMSC-CM protects neurons from apoptosis in vitro. Neuronal death and apoptosis rapidly followed by oligodendrocyte apoptosis are parts of secondary processes making the lesion worse. BMSC transplantations have been successfully used to Folinic acid calcium salt pentahydrate reduce apoptotic death in the context of SCI, and associated to a better motor recovery. In this study, we demonstrate that BMSC-CM treated spinal cords have a reduced depth of cystic cavity, protecting white 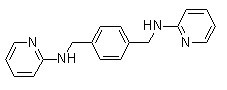 matter tracts. You et al. demonstrated a positive correlation between spared ventral white matter and the final BBB scores of rats. Also, the rubrospinal tract in the dorsolateral part and the corticospinal tract located in the dorsal part of the spinal cord white matter in rats are particularly important for precise limb movements, and can be assessed by grid navigation test. Based on our behavioural data, we can thus conclude that the better motor recovery observed in BMSC-CM treated animals is a direct consequence of improved tissue sparing. Among factors identified in BMSC-CM by cytokine arrays and ELISA, some may also contribute to tissue preservation. NGF stimulates the survival of sympathetic and sensory neurons, while TIMP-1 and CINC-3 are neuroprotective. BDNF administration decreases apoptosis and demyelination in a spinal cord compression model and reduces astroglial scar formation. Also, other factors, thus not described here, are known to be secreted by rat BMSCs: IGF-1, HGF, TGF-b1, EGF, SDF-1, MIP-1a/b, GM-CSF or FGF-2. The fact that we didn’t find those factors is first due to the fact that some of them were not included in our 90-protein array assay. Moreover, BMSC culture conditions may influence their secretome, which would explain why factors described in other studies were not detected here and conversely. In our model, BMSC-CM doesn’t affect astroglial reactivity. This result could be considered as unexpected, as both NGF and BDNF, present in BMSC-CM, are known to reduce reactive astrogliosis. This discrepancy is likely due to the variable concentrations of these two neurotrophins. Also, according to the literature, few studies report any effect of BMSC transplantation on astroglial scar development after SCI. This is also the case for axonal regeneration, which is rarely reported as associated to improved recovery after BMSC transplants. In vitro data on IFNc/LPS-activated macrophages show that BMSC-CM further favours their pro-inflammatory state, as assessed by their Diacerein significant increased IL-1b secretion and their obvious but non-significant increased IL-6 and TNFa production. In parallel, we also show that BMSC-CM contains IL-6, which possesses pro-inflammatory properties as well. In vivo, we didn’t detect any difference between treated and control groups, in the number of macrophages that invaded the lesion site 1 week post-injury, as assessed by the total area stained for CD11b. Microglia/macrophages present within the lesion exhibited in both groups round cell bodies without branching processes, characteristic of “amoeboid” cells, and indicative of an activated status.
matter tracts. You et al. demonstrated a positive correlation between spared ventral white matter and the final BBB scores of rats. Also, the rubrospinal tract in the dorsolateral part and the corticospinal tract located in the dorsal part of the spinal cord white matter in rats are particularly important for precise limb movements, and can be assessed by grid navigation test. Based on our behavioural data, we can thus conclude that the better motor recovery observed in BMSC-CM treated animals is a direct consequence of improved tissue sparing. Among factors identified in BMSC-CM by cytokine arrays and ELISA, some may also contribute to tissue preservation. NGF stimulates the survival of sympathetic and sensory neurons, while TIMP-1 and CINC-3 are neuroprotective. BDNF administration decreases apoptosis and demyelination in a spinal cord compression model and reduces astroglial scar formation. Also, other factors, thus not described here, are known to be secreted by rat BMSCs: IGF-1, HGF, TGF-b1, EGF, SDF-1, MIP-1a/b, GM-CSF or FGF-2. The fact that we didn’t find those factors is first due to the fact that some of them were not included in our 90-protein array assay. Moreover, BMSC culture conditions may influence their secretome, which would explain why factors described in other studies were not detected here and conversely. In our model, BMSC-CM doesn’t affect astroglial reactivity. This result could be considered as unexpected, as both NGF and BDNF, present in BMSC-CM, are known to reduce reactive astrogliosis. This discrepancy is likely due to the variable concentrations of these two neurotrophins. Also, according to the literature, few studies report any effect of BMSC transplantation on astroglial scar development after SCI. This is also the case for axonal regeneration, which is rarely reported as associated to improved recovery after BMSC transplants. In vitro data on IFNc/LPS-activated macrophages show that BMSC-CM further favours their pro-inflammatory state, as assessed by their Diacerein significant increased IL-1b secretion and their obvious but non-significant increased IL-6 and TNFa production. In parallel, we also show that BMSC-CM contains IL-6, which possesses pro-inflammatory properties as well. In vivo, we didn’t detect any difference between treated and control groups, in the number of macrophages that invaded the lesion site 1 week post-injury, as assessed by the total area stained for CD11b. Microglia/macrophages present within the lesion exhibited in both groups round cell bodies without branching processes, characteristic of “amoeboid” cells, and indicative of an activated status.